.jpg "Реализованные проекты")


Медиатека
Многоуровневая эпигенетика: множественные взаимодействия в функционировании репрессоров Polycomb
Статья Александры Головниной, студентки магистерской программы «Структурная биология и биотехнология» МГУ имени М. В. Ломоносова и стипендиата Компании Хеликон.
Компания Хеликон всегда стремилась поддерживать отечественных учёных – как опытных специалистов, так и студентов, которые только делают первые шаги в науку. С этой целью мы учредили ежегодную стипендию «Хеликон» для молодых биологов со всей страны. Первыми стипендиатами стали студенты магистерской программы «Структурная биология и биотехнология», реализуемой на базе МГУ имени М.В. Ломоносова.
Сегодня мы бы хотели поделиться с вами статьёй Александры Головниной. В ней рассказывается о роли группы репрессоров Polycomb в транскрипции многоклеточных организмов.
Хорошо известно, что транскрипция (прочтение закодированной в ДНК информации) – ключевой этап генной экспрессии. Исследования последних десятилетий показали, что в живом организме этот процесс часто не работает по принципу простого линейного ответа на стимул. В одних случаях он должен быстро включаться и выключаться в зависимости от стадии развития, типа ткани и геномного контекста; в других случаях – настраиваться более тонко. Для обеспечения корректного и своевременного транскрипционного ответа клетка задействует множество систем контроля, которые изменяют структурное и функциональное состояние хроматина (комплекса ДНК и регуляторных белков).
Одна из таких систем представлена группой репрессоров Polycomb (PcG), консервативной среди многоклеточных. 1 Гены, кодирующие эти белки, открыли в 1940-х годах у мушки дрозофилы: при мутации в некоторых из них у самцов появлялись дополнительные ряды характерных щетинок – половые гребни (sex combs) – на средних и задних конечностях, в то время как в норме эти гребни должны находиться только на передних конечностях. Тогда учёные предположили, что гены данной группы отвечают за специфическое развитие конечностей. Только почти 30 лет спустя произошёл концептуальный прорыв: при детальном изучении PcG-мутантов у личинок выявили широкий спектр тонких нарушений в формировании передне-задней оси тела и сегментации. Представители группы Polycomb оказалась важнейшими регуляторами онтогенеза. Основная их функция состоит в поддержании репрессии генов, которые были активны в эмбрионе – так они отвечают за сохранение программы, запущенной на начальных этапах развития.
Последующие биохимические исследования показали, что белки PcG функционируют в виде нескольких мультимерных комплексов, основные среди которых – PRC1 и PRC2 (Polycomb repressive complex 1 и 2). Они могут как непосредственно комактизировать хроматин, так и опосредованно, тормозя инициацию и элонгацию транскрипции. Кроме того, PRC1 и PRC2 проявляют активность эпигенетических «читателей» и «писателей», то есть вносят и распознают определённые химические модификации в белках-гистонах, упаковывающих ДНК. Так, метка H3K27Me3 (то есть триметилирование 27 лизина третьего гистона) специфична для Polycomb-зависимой репрессии. Привлечение PcG-комплексов сопровождается обособлением обширных областей хроматина, перестройкой его пространственной структуры и переводом в неактивное, то есть не транскрибируемое состояние. Согласно последним данным, белки Polycomb отвечают не только за импринтинг генома, но также вовлечены в другие фундаментальные клеточные процессы, такие как поддержание стволовых клеток, контроль клеточного цикла и инактивация Х-хромосомы. 2 При этом некоторые из своих функций они выполняют индивидуально, а не в составе репрессорных комплексов.
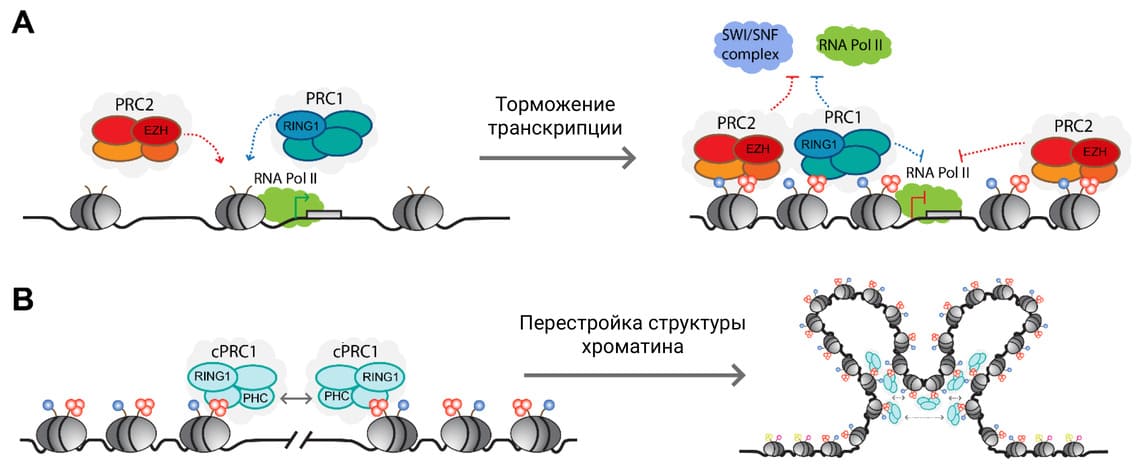
Рисунок 1. Формирование репрессированного Polycomb-зависимого хроматина у человека.
3 А. PRC1 и PRC2 – комплексы PcG, EZH (Enhancer of Zeste homolog) и RING1 – каталитические субъединицы в их составе, вносящие гистоновые модификации (обозначены синими и оранжевыми кружками) в октамеры гистонов, связывающие ДНК – нуклеосомы (обозначены серыми овалами). Стрелками показано их связывание с активным хроматином и репрессивные эффекты на транскрипцию (SWI/SNF – комплекс-ремоделер хроматина). B. Субъединица PHC в составе PRC1 обеспечивает дальние контакты между комплексами PRC1, что способствует перестройке и обособлению репрессированной Polycomb-зависимой области хроматина.
Несмотря на активное изучение системы Polycomb, в деталях её работы многое остаётся неизвестным. В частности, нет однозначного понимания механизма привлечения белков PcG на хроматин. 4 В то же время, это важное направление исследований. Знание механизмов рекрутирования может помочь точечно изменять или исправлять дисрегуляцию Polycomb-репрессии в конкретных геномных областях на уровне связывания комплексов с хроматином, что было бы удобно, так как это не нарушало бы других функций PcG-белков.
Коровые субъединицы Polycomb не связывают ДНК – соответственно, они нуждаются в рекрутерах. Накопленные данные указывают на участие РНК, гистоновых модификаций и транскрипционных факторов в формировании PcG-зависимого хроматина, но предложенные для них механизмы пока не складываются в единую картину. Скорее всего, для правильной регуляции генов необходимо использование нескольких перекрывающихся способов рекрутирования. Согласно одной из моделей, для устойчивого нацеливания PcG-комплексов на регуляторные элементы нужны специфические ДНК-связывающие белки, и это связывание служит точкой нуклеации для дальнейшего распространения, усиления и поддержания PcG-опосредованной репрессии, которое обеспечивается РНК и гистоновыми модификациями. 5
Благодаря консервативности системы Polycomb, дрозофила является удобным модельным организмом для изучения механизмов рекрутирования. У плодовой мушки комплексы PcG привлекаются на специальные регуляторные элементы – PRE (Polycomb response elements), представляющие собой участки ДНК протяжённостью в сотни пар нуклеотидов, плотно «усеянные» множеством сайтов ДНК-связывающих белков. 4 Для каждого PRE количество и взаиморасположение этих сайтов отличается и образует площадку для посадки уникального набора транскрипционных факторов, а эта «мозаика», в свою очередь, определяет уникальный набор связывающихся регуляторных комплексов, а следовательно – характер и силу репрессии.
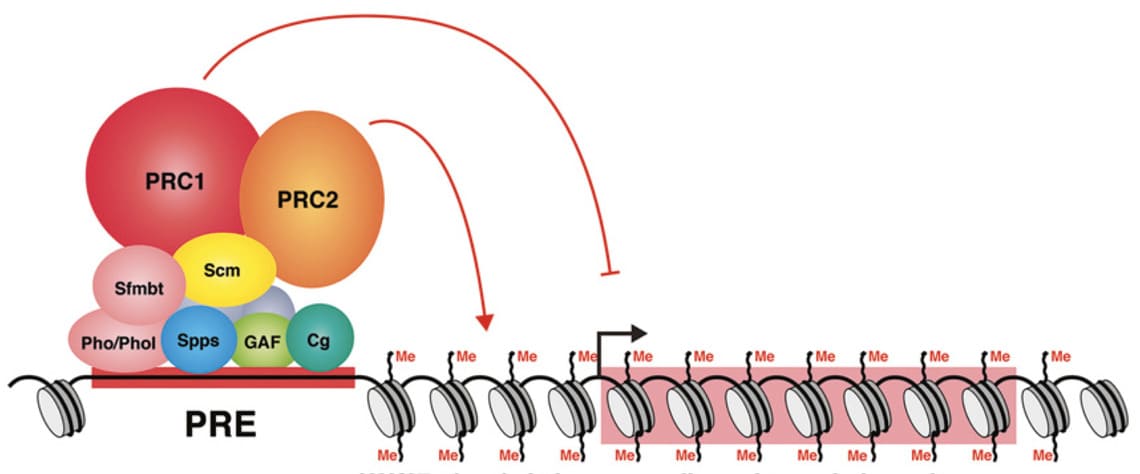
Рисунок 2. Комплексы PRC1 и PRC2 на PRE.
1 Стрелками показана активность по модификации и репрессированию хроматина. Sfmbt и Scm – некоторые адаптерные белки группы Polycomb; Pho, Phol, Spps, GAF и Cg – некоторые PRE-ДНК-связывающие белки. Нуклеосомы обозначены серыми овалами, триметилирование гистонов H3K27 отмечено как Me, красным широким прямоугольником выделен репрессированный ген.
При этом ни одного ДНК-связывающего белка в отдельности недостаточно для надёжного привлечения PcG-белков и возникновения репрессии – необходимы кооперативные взаимодействия между рекрутерами. Такие сложные паттерны контактов могут быть необходимы для устойчивости и, в то же время, возможности тонкой настройки Polycomb-опосредованной репрессии. Кроме того, множественные взаимодействия обеспечивают специфичность привлечения PcG-белков, так как все PRE-ДНК-связывающие факторы являются полифункциональными и участвуют не только в репрессии транскрипции, но и в её активации на разных регуляторных элементах. Более того, некоторые PRE способны эпигенетически переключаться и на других стадиях развития поддерживать транскрипцию генов-мишеней – в этом случае часть из этих же ДНК-связывающих беков обеспечивает привлечение на PRE активирующих комплексов группы Trithorax. В таких регуляторных элементах итоговый эффект на гены-мишени определяется соотношением активирующих и репрессорных комплексов.
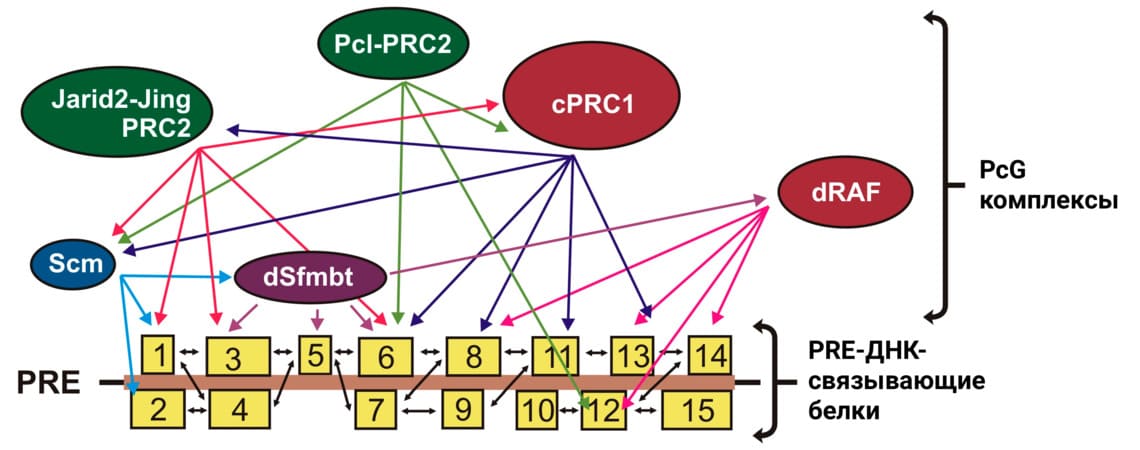
Рисунок 3. Модель комбинаторного рекрутирования системы Polycomb на PRE. Множественные ДНК-связывающие факторы (обозначены жёлтыми прямоугольниками 1-15) взаимодействуют друг с другом и с разными белками PcG, создавая специфическую платформу, эффективную для привлечения определённого набора PcG-комплексов.
4
На данный момент известно более 10 PRE-ДНК-связывающих факторов. Предполагается, что их ещё больше. Однако возможные механизмы действия определены только для некоторых из них. GAF (GAGA factor) и Grh (Grainy head), вероятно, действуют как «пионерные факторы»: они связываются с «закрытым» хроматином, недоступным для большинства факторов транскрипции из-за высокой нуклеосомной плотности, локально его декомпактизируют и этим облегчают привлечение остальных регуляторов 6, 7. В выявлении индивидуальных ролей PRE-ДНК-связывающих белков помогает протеомика, в частности анализ их интерактомов, то есть совокупности прямых и опосредованных взаимодействий. Недавние исследования показали, что у отдельных PRE-ДНК-связывающих белков интерактомы в разной степени обогащены представителями разных регуляторных систем 8. Например, факторы Combgap и Zeste наиболее сильно ассоциированы с комплексами Polycomb, тогда как Psq (Pipsqueak) – с активаторами группы Trithorax, конкурирующими с системой Polycomb на PRE, и ремоделерами хроматина. Интерактом Adf1 (Adh transcription factor 1) наиболее обогащен компонентами медиаторного комплекса – важнейшего компонента транскрипционной машинерии. Некоторые также ассоциированы с промоторными факторами и белками, поддерживающими архитектуру хроматина. Также замечено глобальное подразделение PRE-ДНК-связывающих факторов на две подгруппы в соответствии с их способностью к взаимодействию между собой и с субъединицами одних и тех же регуляторных комплексов. Одна подгруппа функционально близких и более плотно контактирующих белков включает Pho, Spps, Phol и Grh, другая – Combgap, Zeste, Psq, Adf1 и Dsp1.
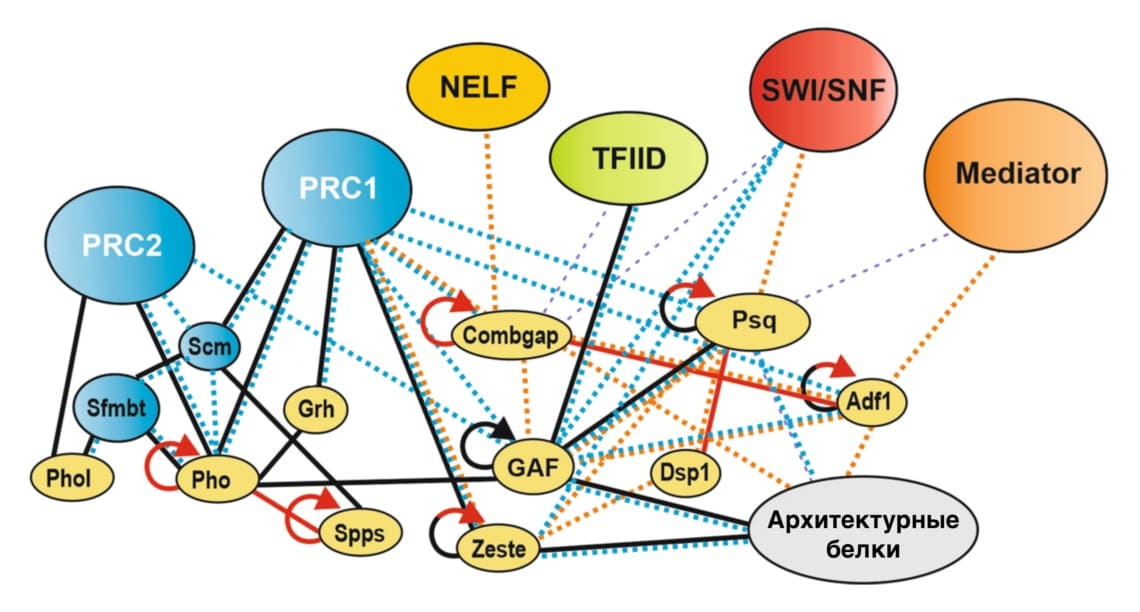
Рисунок 4. PRE-ДНК-связывающие факторы (окрашены жёлтым) взаимодействуют с компонентами различных регуляторных комплексов. PRC1 и PRC2 являются комплексами системы Polycomb; TFIID и NELF участвуют, соответственно, в инициации и запуске продуктивной элонгации транскрипции; Медиатор относится к машинерии РНК-полимеразы; SWI/SNF – комплексы ремоделеров хроматина. Физические контакты, установленные непрямыми методами, обозначены пунктирными оранжевыми линиями (сильное взаимодействие) или фиолетовыми (слабое взаимодействие); прямые партнеры обозначены сплошными линиями. Способность белков к гомоолигомеризации показана круглыми стрелками. Красные линии, оранжевый и фиолетовый пунктир отражают данные, полученные на основе подробного интерактомного анализа.
8
Детальное изучение обнаруженных функциональных связей и выявление прямых партнёров PRE-ДНК-связывающих факторов должно помочь в решении головоломки о множественных взаимодействиях на PRE и восстановлении порядка событий, происходящих на при рекрутировании Polycomb-комплексов.
Автор: Головнина Александра
Литература
1. Kassis J.A., Kennison J.A., Tamkun J.W. Polycomb and Trithorax Group Genes in Drosophila // Genetics. – 2017. – V. 206. – № 4. – P. 1699-1725.
2. Schuettengruber B., Bourbon H.M., Di Croce L., Cavalli G. Genome Regulation by Polycomb and Trithorax: 70 Years and Counting // Cell. – 2017. – V. 171. – № 1. – P. 34-57.
3. Flora P., Dalal G., Cohen I., Ezhkova E. Polycomb Repressive Complex(es) and Their Role in Adult Stem Cells // Genes (Basel). – 2021. – V. 12. – № 10.
4. Erokhin M., Georgiev P., Chetverina D. Drosophila DNA-Binding Proteins in Polycomb Repression // Epigenomes. – 2018. – V. 2. – P. 1.
5. Cheutin T., Cavalli G. The multiscale effects of polycomb mechanisms on 3D chromatin folding // Crit Rev Biochem Mol Biol. – 2019. – V. 54. – № 5. – P. 399-417.
6. Chetverina D., Erokhin M., Schedl P. GAGA factor: a multifunctional pioneering chromatin protein // Cell Mol Life Sci. – 2021. – V. 78. – № 9. – P. 4125-4141.
7. Sundararajan V., Pang Q.Y., Choolani M., Huang R.Y. Spotlight on the Granules (Grainyhead-Like Proteins) - From an Evolutionary Conserved Controller of Epithelial Trait to Pioneering the Chromatin Landscape // Front Mol Biosci. – 2020. – V. 7. – P. 213.
8. Chetverina D., Vorobyeva N.E., Mazina M.Y., Fab L.V., Lomaev D., Golovnina A., Mogila V., Georgiev P., Ziganshin R.H., Erokhin M. Comparative interactome analysis of the PRE DNA-binding factors: purification of the Combgap-, Zeste-, Psq-, and Adf1-associated proteins // Cell Mol Life Sci. – 2022. – V. 79. – № 7. – P. 353.